(注)手書きの卒業論文を部分的に入力。実物は、製本され、大学の図書室に保管されている。→【卒業論文(PDFファイル)】
(注)卒業論文の参考文献の調査メモが50年ぶりに見つかった。→【参考文献の調査メモ(PDFファイル)】
卒業論文 :条件反射の生体工学的解析
1969年5月12日提出
指導教官 斎藤正男 助教授
東京大学工学部電子工学科
中所武司
序文
我々、人間の肉体に似ていて、道徳的にも可能な限り、我々の行為をまねる機械があるとしても、我々はやはりこれらの機械が、だからといって真の人間ではないことを認識する二つの極めて確かな方法を持っている。
(1637 デカルト)
人間はきわめて複雑な機械である。一挙にして明らかなる観念を持つことは不可能であり、従ってこれを定義することは不可能である。最も偉大な哲学者たちがアプリオリに,すなわち,いわば精神の翼の力を借らんとして、なしたすべての探究がむなしかったのは正しくこのためである。かくして,アポステリオリに、すなわち、いわば、人体の諸器官を通して、霊魂の姿を見わけようと試みることによって初めて、人間の本質そのものを明確に発見できるとは言わないが、この点に関して可能なる限り、最高度の蓋然性に到達しうるのである。
(1747 ド・ラ・メトリ)
手と言語器官と脳の協同作業によって、各人にあってのみならず、社会の中でも、人間はますます複雑になっていく諸作業を遂行し、いよいよ高い諸々の目標を自らに課し、かつそれを達成することができるようになった。
(1876 エンゲルス)
今年は1969年である。
今から約300年前、動物は機械であると言ったデカルトも、人間の脳を前にしては、神の助けを借りないわけにはいかなかった。
今から約200年前、フランスの医師ド・ラ・メトリは人間は機械であると考えたが、それを実証しようとするには、あまりに絶望的になっていた。彼は結論を急ぎすぎたのだ。
今から約100年前、かのエンゲルスも、人間を外からしか見ようとはしなかった。
それでは、現在、我々は脳についてどれだけのことを知っているであろうか。脳生理学は、確かに100年前と比べるべくもないが、まだまだ幼児期であり、今は学問としての確立期のように思われる。脳に対するアプローチの仕方は、まだ暗中模索であり、それだけに学者間の考えの違いは著しく、欧米系とソ連系の間でも、大きな対立があるような状況である。現在は、電気生理学、条件反射学,解剖学など、あらゆる分野で多くの実験が行われ、幾多の事実が明るみに出されている段階で、それらをもとにした、まとまった理論的考察は皆無に等しい。
一方、生体の持つ、情報処理、学習機能についての工学分野からの興味は尽きない。
それで私は、生物の持つ学習機能、特に、その最も単純化されたもの、あるいは最も基本的なものと思われる条件反射を取り上げて脳の機能を探ってみようと思う。
目次
序文
第1章 条件反射の生理学
1.1 条件反射
1.2 脳の働き
1.3 神経系と神経細胞
第2章 条件反射の生体工学的解析
2.1 条件反射の学習性
2.2 記憶
2.3 シナプス結合による条件反射の解析
2.4 この方法論の限界と克服
第3章 学習機械としての、条件反射を行う脳のモデル
3.1 神経回路網の機能の分解と合成
3.2 組み合わせモデル
3.3 集積回路化について
3.4 学習機械としての評価
第4章 結言
参考文献
第1章 条件反射の生理学
1.1 条件反射
動物は生まれながらに幾多の能力を備えている。そのうち、あるものは無条件反射と呼ばれる。それはある入力信号に対しては、決まってそれに対応するある出力を出す反応であり、食餌を口に入れると唾液を分泌したり、あるいは、肢に電気刺激を与えるとその肢を屈曲させたりする反応のことである。動物たちは、これら無条件反射を基礎として、その上に生活の中でのいろいろな経験を積み重ねていくのであり、それは条件反射の積み重ねである。初めて肉を見た犬は、それだけでは唾液を分泌しないが、それが美味しい食餌であることを知ると、その後は肉の匂いだけで唾液を分泌するようになる。このように自然に経験したものは自然条件反射という。また、パブロフの実験で有名だが、犬に食餌を与える前にベルを鳴らすことを繰り返すと、犬はベルの音を聞いただけで唾液を分泌するようになる。このような反応を人工条件反射と言う。
ひとたび、ベルで食餌条件反射を形成された犬は、いつまでもベルさえ鳴れば唾液を分泌することを繰り返すかと言うとそうではない。ベルの音の後で、食事を与えて強化しないと、条件反射は消えてしまうのである。この起こるべき条件反射が起こらないという抑制作用が、動物の生存に重要な働きをしている。そこで、次に条件反射の抑制作用について述べる。
抑制には、大きく分けて外抑制(無条件抑制)と内抑制(条件抑制)とがあり、前者は外部的要因によるもので、危険を感じた時に起こす防御反応のために起こる抑制(恒常抑制)や、新奇な信号に対して起こす定位反射(詮索反射とも言う)のためにおこる抑制(減衰抑制)などがあり、内抑制では主なものは四つで、そのひとつは先に述べたように、強化しなければ徐々に条件反射反応が弱まっていく消去抑制である。これは条件反射の完全な消滅ではないので、消去後に再び強化すればすぐに回復するのである。この抑制は、訓練されるのであって、もし条件反射の消去と反射を繰り返せば、最後には、一度強化なしの条件刺激を与えただけで、条件反射は消去されるようになる。
第二には、分化抑制があり、これはメトロノームを条件刺激に使って、毎分60 bit の時は食餌を与え、毎分120 bitの時は与えないようにして何度か訓練すると、信号の分化が起こり、毎分60ビットでは唾液を分泌するが、毎分120ビットでは分泌しなくなるような反応である。この場合、陽性信号(興奮を引き起こす信号)に近い陰性信号(抑制を引き起こす信号)に対する分化抑制は困難である。メトロノームで毎分60 bit と毎分120bitの分化は40ないし50回の訓練で完成するが毎分60 Bit と毎分70bitのときは100ないし200回必要で、それでも分化できない犬もいる。訓練をする時、陽性信号と陰性信号を交互に与えてやると、早く分化が形成され、また毎分60 bit と毎分120 Bit の分化を形成させた後に、陰性信号を120ビットから70ビットまで徐々に変化させてやると、はじめは分化できなかった犬でも分化ができるようになる。
第3には、付加抑制があり、これはメトロノームで食餌条件反射を形成後、メトロノームと同時に水泡音を付加した時には食餌を与えないことを繰り返すと、メトロノームだけの時は唾液を分泌するかそれに水泡音が加わると唾液を分泌しなくなる反応である。この抑制の形成には付加刺激と条件刺激との強さの相対的な関係が重要な要因となっており、付加信号が強いほど、また陽性信号が弱いほど形成が容易である。最後に延滞抑制があり、これはベルを鳴らした後5分間経過すると食餌を与えることを繰り返すと、ベルの音を聞いてから5分後に唾液が分泌されるようになる反応である。この抑制では、時間がひとつの信号になっており、陽性信号の強いほど、また反射の強さの大きいものほど、形成が困難である。以上のような多様な抑制作用が、動物のより複雑な行動を可能にしているのである。
この抑制作用には相互作用があり、この作用による新たな反応もまた重要である。それはある抑制過程が他の抑制過程を弱めたり消したりする条件反射の脱抑制、すなわち抑制が解けて条件反応を起こしたり、逆に一方が他方を強めるような抑制の加重であったりする。例えばメトロノームで形成した食餌条件反射を完全に消去した後、メトロノームと同時に口笛を付加すると、脱抑制して唾液を分泌するようになったり、ある音を陽性信号、その1/8音を陰性信号として分化抑制を完成させた後、1/8音と同時に水泡音を付加すると脱抑制して唾液を分泌するようになる。また延滞抑制の条件反射を形成後の消去は、普通の条件反射(陽性信号と強化の間が1ないし2秒)の消去よりも速やかに形成されるという抑制の加重も見られる。
以上、動物の、特に犬を例にしながら、正常状態での条件反射および条件抑制について概観してきたが、その他に病的条件反射と言われているものがある。例えば0.05%アコニチン1ml の投与で不整脈を伴う頻脈を生じるという無条件反射を使って、これと条件刺激の音とを組み合わせると音だけで頻脈を生じる。大量のカルボコリンで狭心症を起こさせ、これと音を組み合わせると、音だけで発作が起こる。ある指揮者がある曲で失敗して発作が起こるという事件のあと、その曲の時はいつも発作起こしてしまうという例はこの場合にあたる。これと同じような事は日常生活でもよくあることで ある。これら病的条件反射の特徴は、正常な条件反射では例えば食餌条件反射性唾液分泌は無条件反射性分泌の半分以下であるのに、病的条件反射ではほとんど無条件反射に近い強さで反応があらわれることである。
今までに述べてきた条件反射の形成法は、古典的条件付けとか、応答的条件付けとか呼ばれ、パブロフによって導入され、それ依頼使われている方法であるが、それに対応するものとして、道具的条件付けとか、操作的条件付けとか言われるものがある。例えばレバーを押せば食餌が与えられる仕掛けのスキナー箱の中にネズミを入れると、ネズミは食餌を得るために連続的に速くレバーを押すようになるというような報酬訓練とか、一つの部屋を二つに仕切り、一方は壁を白く塗って床から電気刺激を与え、もう一方は壁を黒く塗って木の底にしておくと、白い部屋に入れられたネズミは全速力で黒い方へ逃げるようになるという避難訓練とか、モルモットを回転籠に入れブザーを鳴らして2秒後に回転籠に電気刺激を与えるようにし、もし回転籠をある角度以上回せば このショックを回避できるようにしておくと、モルモットはブザーがなるや否や回転籠を回すようになるという回避訓練などが、この操作的条件付けの例である。
この他、条件付けの一種と考えられるものに、図1-1のようなT字迷路を用いて、音信号のあるときは左、ないときは右へ行けば食餌が与えられるようにして、音信号の有無で左右を弁別させる弁別学習とか、正しい道を行けば食餌にありつけるような迷路で正しい道を見つけさせる迷路学習などもある。なおこれら操作的条件付け、弁別学習、迷路学習が古典的条件付けと本質的に異なるものか否かについての考察は後章に譲ることにする。
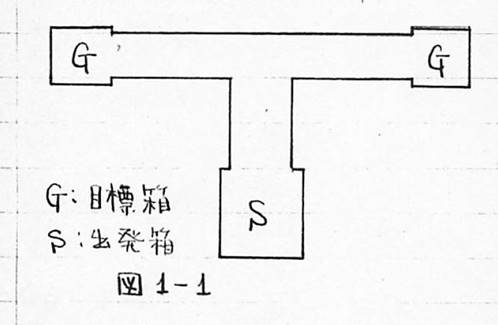
図1-1 ■出発箱1個と目標箱2個をふくむT字型の図■
1.2 脳の働き
前節では、条件反射の現象面を概観してきたのであるが、それでは、いったい、この膨大な入力と出力を持ち複雑な反応を示すアニマルと言われるブラックボックスの中はどうなっているのであろうか。
犬がベルの音を聞いて唾液を分泌するという条件反射反応を考えてみると、環境の変化を感じ取る受容器の一つである耳がベルの音を感受したことと、唾液分泌腺という外界へ働きかける効果器の一つが働いたことは明白である。だから、この受容器と効果器の間で何らかの情報伝達が行われたわけであるか、これには二つの型が考えられる。受容器の電気信号すなわちインパルスをそのまま効果器へ伝える型式と、受容器のインパルスが一度たくさんの神経細胞の集団の中に送り込まれ、複雑な処理をされた後、効果器へ適当な信号が送られるものがある。前者の場合には、神経細胞は単なる伝導器の働きをしており、単純な無条件反射はこの場合である。後者は神経回路網が統合作用を行っており、犬の食餌条件反射はこれにあたる。
受容器と効果器について簡単に述べておくと、受容器は色々な形で吸収される外界のエネルギーを電気エネルギーに変えるトランスジューサーの役割を果たしており、それには機械ー電気系、熱ー電気系、化学ー電気系、光ー電気系などがある。効果器には筋肉と分泌腺がある。受容器の感覚信号としてのインパルスは、2ないし5個の中継核で中継された後、新皮質のそれぞれの感覚野に投射している。この新皮質の運動野から出たインパルスは、それぞれの運動神経を通って筋活動を行う。しかし、情報は決して一方向性ではなく、常にフィードバックがあることが分かっており、そのために感覚器、効果器が適切な働きをしうるのである 。
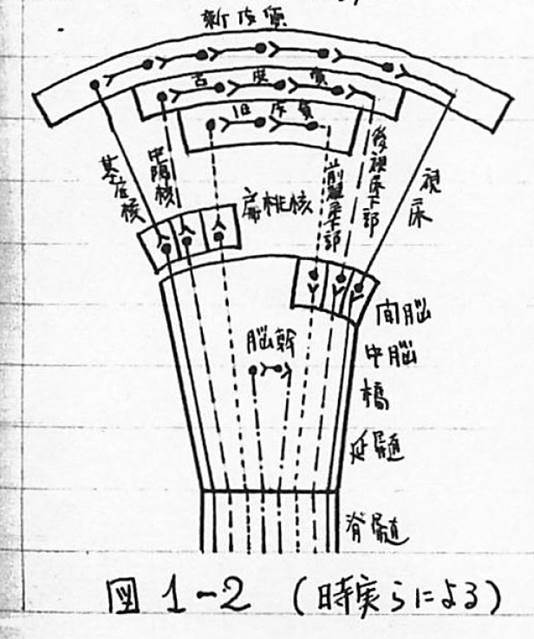
図1-2 (時実らによる)■新皮質から脊髄に至る詳細図■
次に条件反射活動の中枢部にあたる脳の働きについて述べる。脳で統合作用を受けもつものには、新皮質系、古皮質系、旧皮質系、脳幹系などがあり、前の3つの求心性神経路が最後に中継する部位は、それぞれ視床、視床下部後部、視床下部前部であり、遠心性神経路が最初に中継する部位は、それぞれ基底核、中隔核、扁桃核である。なお新皮質からの遠心性神経路には基底核で中継されるもののほかに、直接、脊髄へ直行するものがあり、前者を錐体外路、後者を錐体路と言う。これらの統合系の間での役割の分担は決まっており、新皮質系へ行く情報は外部環境の変化であり、光、音の感覚や、皮膚感覚、味覚(甘さ、辛さ)であり、大脳辺縁系(古皮質と旧皮質)へは、内部環境の変化と嗅覚、温覚、圧覚、味覚(苦さ、酸っぱさ)であり、統合作用のうち、最も単純な脳幹脊髄系では、視床下部以外の体内の受容器で受け止められる内部環境の変化と、痛覚、平衡感覚、筋肉の張力の感覚である。図1-2は、これら統合型を模型的に示したものである。
ここで、脳の行う反射活動を考えてみると、無条件反射を司っているのは、大脳半球に最も近い皮質下であり、条件反射は大脳皮質であると考えられている。これは、条件反射の開拓者Pavlov以来の考えであり、彼は、条件反射の形成は、新皮質(大脳皮質も同じこと)での一時的結合であると考えていた。そして、条件反射は常に無条件反射を基礎にして行われることから、皮質下構造と新皮質との相互関係の問題は、脳生理学の一つの大きな基本問題である。
これに関しての興味深い一つの事実は、条件反射の形成過程で、無条件反射が変化することである。食餌条件反射の実験を繰り返し行っていると、唾液分泌の無条件反射が増強されるのである。この現象は、病的条件において、いっそう明確である。毎日、系統的に犬にモルヒネを注射していくと、初めのうちは嘔吐は8分後におこったが、8回目の実験では、それが2分後にまで早められていた。
これについてのソビエトの学者達の考えは、任意の無条件反射が皮質下の準位でばかりではなく、新皮質においても行われるようになり、そのために皮質性インパルスが皮質下性インパルスに加重されるようになるので、無条件反射が改造され、皮質性機能の色合いをだんだん持ってくるというものである。そして、この無条件反射は、いま述べたような皮質下現象のために、より不安定で易動的なものになる。
これらの新皮質の作用を探るために、それを実験的に損傷して、それによる高次神経活動の障害を研究するという解剖学的方法がとられている。その結果、新皮質を除去すると、無条件反射の機能が低下するということがわかった。この事実は、先の無条件反射の皮質下現象という推論とよく合う。また、去脳犬は、すべての自然、及び、人工条件反射を忘れ、何度訓練しても新たに条件反射を形成しなかったというソ連の学者の報告があるが、これは、後に、皮質を除去しても、基本的な条件反射は形成されるという実験データがだされて、修正をうけた。また、側頭葉や後頭葉を切除すると、音は聞こえるが、理解することはできない精神聾や、見えるけれども、それを認識することはできない精神盲を引き起こすというように、皮質分析器の除去は、刺激体の要素的な分析とか総合とかは、手術後、しばらくすると回復するが、細かな分析や総合の障害を起こす。
このことから、ある分析器に属する神経細胞は、大部分が基本的な核のまわりにあるが、一部は、全体に散在していることもわかった。その後、各々の皮質分析器の間を切断した後でも、それらに対応する刺激で条件反射が形成されたことにより、いろいろな皮質分析器相互間の結合は皮質下構造を通じて行われることがわかった。
以上、主にソ連の文献を参考にしながら述べてきたが、欧米には、ソ連系の考え方に批判的な見方をする学者も少なくない。それは、皮質を除去しても、簡単な条件反射は形成されるという、今では通説になっている事実や、欧米で特によく研究されている網様体が非常に重要な機能を持っていることから、学習機能は皮質下にあるとする学者がおり、極端な例では、ソ連では、皮質の機能と考えられているものを網様体の機能であると主張する者もいる。これらの意見の違いは、そのまま、脳生理学がまだまだ初歩の段階にあることを如実に示すものといえよう。
この網様体の研究は、それのある部位を刺激すると、脳波が睡眠パターンから覚醒パターンに移行し、他のある部位を刺激すると、その逆の現象が起こるという実験に始まり、睡眠や意識、覚醒の機序の手掛かりを得るべくおこなわれており、この問題においても、意識の生理学的機序として、いま述べたような網様体調節系を考えるMagounやMoruzziのほかに視床下部調整系を考えるGellhornらがおり、また、時実らによって、この二つの考えを統一的に見ようとする者もあり、まだはっきりは分かっていないが、ここでは、条件反射との関連で網様体の働きを述べる。
以前は、受容器の興奮伝達路は、その対応する皮質感覚野へいく一本だけと考えられていたが、実はもう一本、網様体へいく側枝のあることがわかった。前者を特殊感覚系、後者を非特殊賦活系という。この網様体は、皮質の全面と通路をもっており、受容器の感覚インパルスは、この網様体経由でも、皮質へ伝わるのであって、その時間的遅れは、特殊路に比べて約10msecくらいである。
また、皮質から網様体への働きかけも、その逆の場合に劣らず重要なものである。この網様体は、脳幹部と視床部で働きを異にしており、前者は皮質全体にわたって、徐波の広汎な抑制、すなわち、皮質の持続的、賦活作用を持ち、大脳皮質の活動を全面的に統御しているが、後者は、感覚インパルスを調節して、大脳皮質に局在的に働きかけており、これは、意識の集中や慣れの現象と関連があると思われる。
ここで、光刺激の場合を例として、網様体の働きを述べてみよう。光刺激のインパルスは、はじめ、特殊路と非特殊路の両方に伝わり、前者は、瞳孔収縮という順応反射を引き起こし、後者は、脳幹網様体を興奮させて、皮質のα-リズムの広汎な抑制を行い、同時に、視床部の網様体によって、視覚分析器の皮質投領野に強く作用する。このような効果は、
**<中略>**
ここで、以上の脳の働きを、反射活動に限定してブロック図に表してみると、図1-3のようになる。
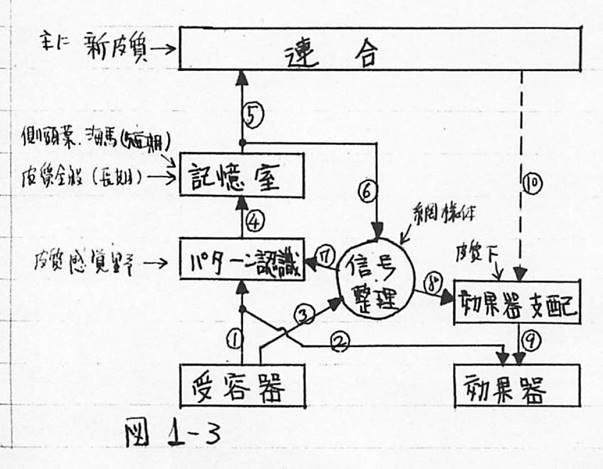
図1-3 反射活動に限定したの脳の働きのブロック図
**<中略>**
1.3 神経系と神経細胞
神経系を形づくるものは、脳膜や豊富な血液網を除けば、神経細胞とグリヤ細胞である。グリヤ細胞は、神経細胞の支柱や、その物質代謝にたずさわるもので、本質的な神経機能は神経細胞が行う。この神経細胞は、生後一年で分裂能力を失い、数は、以後一定である。新皮質だけで140億個と言われている。グリヤ細胞の方は、一生分裂し、数も、神経細胞に比べて、圧倒的に多い。
神経細胞の構造を図1-4に示す。
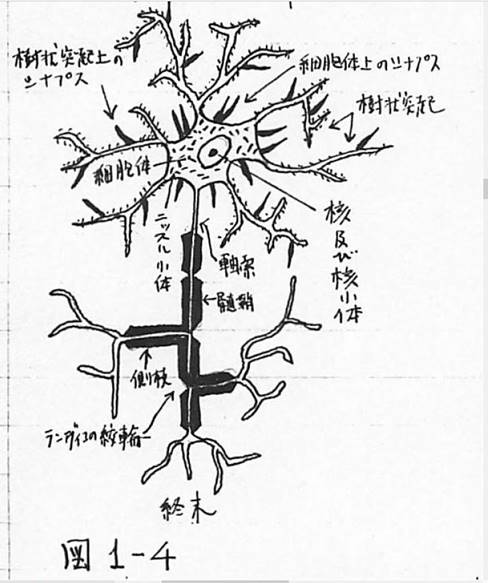
図1-4 神経細胞の構造
細胞体の径は、20〜100μで、形は多様、樹状突起は細胞体から数本〜数十本でるが、枝分かれも多く、これが、他の神経細胞の軸索終端とシナプスをつくる。軸索は長いものは1〜2mあり、1本であるが、ところどころで枝分れして側枝を出す。髄鞘は大部分の神経細胞についており、絶縁物の役割をしている。ところにより、髄鞘がないのは、ランヴィエの絞輪で、軸索の物質代謝をするところらしい。軸索終端は他の神経細胞の細胞体や樹状突起とシナプスをつくる。末梢神経の末端は筋肉細胞、感覚細胞、腺細胞と接触するが、これも一種のシナプスである。そして、これらのシナプスを通して、神経細胞間の興奮の伝達が行われるのである。
神経系の情報伝達はインパルスによっておこなわれ、インパルスの伝導は軸索を通して行われる。軸索を伝導するインパルスを興奮過程と呼び、その本態はイオンである。インパルスは「全か無かの法則」によって伝導し、軸索の径が同じならば、等速度で無損失である。軸索の膜の内側は外側に比べて、静止状態では50〜70mVくらい低電位になっているが、神経細胞が興奮すると、図1-5に示すような、100〜120mVのインパルスが発生し、膜の内側は外側に比べて、40〜50mV高電位になる。
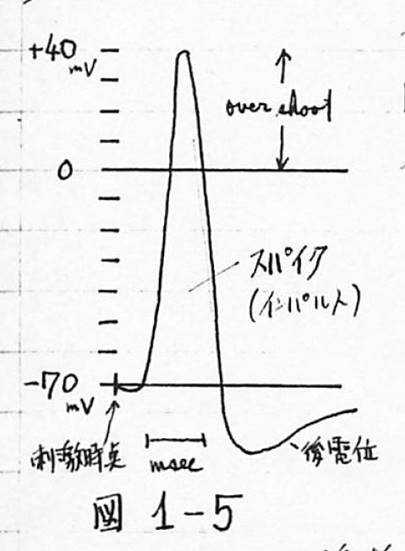
図1-5 神経系の情報伝達を担うインパルス
このインパルスの持続時間は約1msecで、これをスパイク電位といい、つづいて後電位といわれるものがくる。この軸索を通ってきたインパルスは、終末のシナプス小頭部に達する。
シナプスには興奮性シナプスと抑制性シナプスがあり、前者にインパルスが到着すると、細胞膜(シナプス後膜)の電位は脱分極の方向、即ち、正方向に動き、後者の場合は、膜電位は過分極の方向、即ち、負方向に動く。これらのインパルスによる膜電位の変化がある値を越えると、この細胞体は興奮し、スパイクを放出する。この時間は約1msecで、この間は他のインパルスが入ってきても一切反応しないので、絶対不応期と呼ばれ、その後、強い刺激に対してだけ反応する相対不応期があり、つづいて、比較的緩やかな脱分極の時期(10数msec)と長い過分極の時期(約100msec)とがある。
なお、これら神経細胞のシナプス結合を介しての神経系の働きは後章に譲る。
**<中略>**
第2章 条件反射の生体工学的解析
2.1 条件反射の学習性
2.2 記憶
2.3 シナプス結合による条件反射の解析
2.4 この方法論の限界と克服
第2章 条件反射の生体工学的解析
2.1 条件反射の学習性
条件反射の形成において用いられる入力信号としての条件刺激は、多種多様であるが、これらの一つ一つをとって分析してみると、どんな入力信号も、組合せ、順序、時間の3つの基本的要素によって構成されていることがわかる。条件反射及びその分化抑制、付加抑制に用いられる入力信号は、その多くが単純な1つの信号か、或いは、いくつかの単純な信号の組合せである。また、犬は、音―カサルカー光と、光―カサルカー音とを、その順序の違いによって分化できるが、これは、入力信号が、組合せと順序でできている。
時間の例としては、延滞抑制や、その他にメトロノームで100bits/分と96bits/分とを犬は聞き分けて分化することができることなどがある。但し、ここで考えている時間というのは、延滞抑制の場合で数分、メトロノームなどの場合で1秒前後の時間間隔である。結局、条件反射活動は、上に述べた3つの基本要素からなる入力信号の一つに対して、ある特定の出力を対応付けることと考えていいだろう。
これが,そのまま条件反射の学習機能であると考えても不都合はないが、もう少し詳しく分析してみる。私は序文で条件反射は学習の最も単純なもの、あるいは最も基本的なものであると述べたけれども、それでは条件反射のどこを持って学習と考えるべきであろうか。
**<中略>**
条件反射に特有なものは、連合操作だけであり、その他のパターン認識とか、そのパターンの記憶などは、一般的なものである。
**<中略>**
さて、ここで、1.1で述べた道具的条件付け、弁別学習、迷路学習などの学習過程を、古典的条件付けとの比較で検討しておく。これら3つの学習過程に共通することは、古典的条件付けと違って環境に対する積極的な働きかけのあることである。
**<中略>**
結局、古典的条件反射も道具的条件付けも学習過程の本質は同じである。
2.2 記憶
2.1からわかるように、条件反射の形成において、必要する記憶は、2種類ある。一つは、入力信号の記憶であり、今一つは、ある入力とある出力との対応の記憶である。
**<中略>**
さて、この記憶をつかさどる脳の部位は、一部は新皮質の側頭葉、古皮質の海馬であることが分かっている。子の側頭葉や海馬を除去したサルやラットでは、強い記憶障害がおこり、学習の形成が極めて悪くなり、定位反射はいつまでも消えない。しかし、以前に形成された学習はそのまま残っている。
また、ラットの両側の海馬に拡延性抑制をおこさせると、条件反射はほとんど完全に消失した。
また、精神病の治療のために、側頭葉を外科的に切除した患者は、新しい経験を記憶できず、手術前2〜4年の記憶は消えるが、古い記憶は十分想起できるのである。
**<中略>**
とにかく、記憶をその部位が受け持つにせよ、記憶の前後で何らかの変化が起こるはずだが、それは何であろうか。古くからの考えとしては、神経回路の形成があり、何か刺激があると、そのインパルスによって、神経の閉回路網ができ、インパルスが保持されるというような反響回路を考えて、これを一時的記憶として、一方、このインパルスの持続過程でシナプスに構造的変化が起こることが長期の記憶であるとする考えがある。
このような考えは今でも健在であり、学習過程における生体内の変化は、シナプスにおける何らかの変化であるとして、具体的には、シナプスにおける抵抗の減少、即ち、インパルスを伝えやすくなることが学習の本質であるとする。
この機構については、主な考えに三つあり、一つには、インパルスがシナプスを何度も通ると、シナプス桿状体が膨大になり、伝導度が高くなるという解剖学的考え、第二には、シナプスにおける伝導物質としてのアセチルコリンが増大して、伝導度が高くなるという考え、第三には、グリヤ細胞との関連で、説明しようとする試みなどであるが、明らかではない。前の二つについては、一応の実験的裏付けがある。
しかし、最近、学習による変化をシナプス以外に求めることが試みられている。それは、生化学的変化を考えて、学習における変化は、細胞質を組み立てているたんぱく質の構造的変化とする。即ち、RNA(リボ核酸)と記憶との関連を主張するのだが、それに関するデータとしては、神経細胞を刺激すると中のRNAが増加し、ほっておくと減少することや、RNAを補給すると老人の記憶力の回復、或いは、ネズミの学習能力の向上が起こること、光に対して体を曲げる反応を学習した頭部と尾部に折半してそのまま再生すれば、共に学習を保持しているが、RNA分解酵素のリボヌクレアーゼの中で再生させると、尾部の方は学習が消えたこと、或いは、学習したプラナリアを食べたプラナリアは、その学習を受け継ぐことなどが分かっている。
しかし、一方、RNAと学習との関連に否定的な実験結果もあり、今は明らかではない。
2.3節 シナプス結合による条件反射の解析
**<中略>**
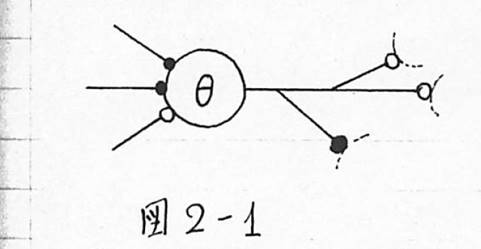
図2-1 McCulloch-Pittsの神経回路網のモデル
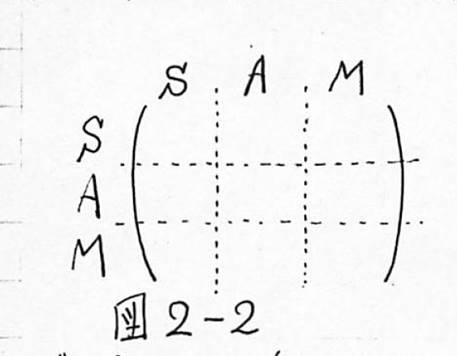
図2-2 Harthの結合マトリックスのモデル
(S:感覚神経系、A:連合野の神経系、M:効果器の神経系)
そこで,このHarthやCaianielloのモデルの限界を考えてみると,彼らの考えた神経細胞の学習効果は第iと第jの神経細胞が興奮すればその間の結合係数が増加するというものであったが,この場合には,もし神経系の中にあらゆる他の神経細胞との結合係数が0であるような神経細胞があっても,これは永久に孤立したままで,使用されることがない.このような神経細胞を以後は自由な神経細胞と呼ぶことにすることにする.こういうことが起こりうるということは,逆に考えれば,学習効果は既に入力から出力までのルートの完成している神経細胞と入力の神経細胞の間にしか現れないことになり,Harthのモデルで最後の形の条件反射が形成不可能であったのは このためである.やはり,このような条件反射,即ち非線形なものの分離をやらせるためには自己組織系の中に先に述べた自由な神経細胞の加わってくる必要があると思われる.なお,この理論的な考察は次節に譲る.
2.4 この方法論の限界と克服
**<中略>**
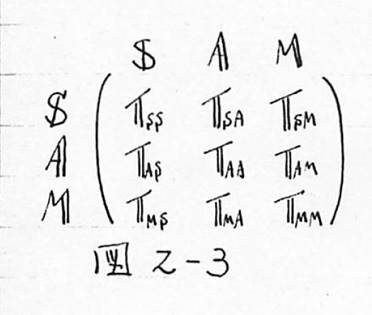
図2-3 Harthの結合マトリックスのモデル(区分別の行列の表示)
第3章 学習機械としての、条件反射を行う脳のモデル
3.1 神経回路網の機能の分解と合成
3.2 組み合わせモデル
3.3 集積回路化について
3.4 学習機械としての評価
第3章 学習機械としての、条件反射を行う脳のモデル
3.1 神経回路網の機能の分解と合成
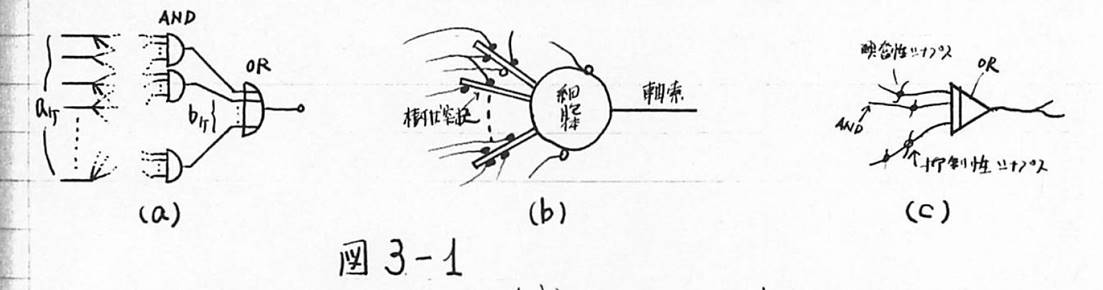
図3-1 神経細胞の回路表現
(a)神経細胞の論理素子表現 (b)樹状突起がAND回路、細胞体がOR回路 (c)神経細胞の図式化
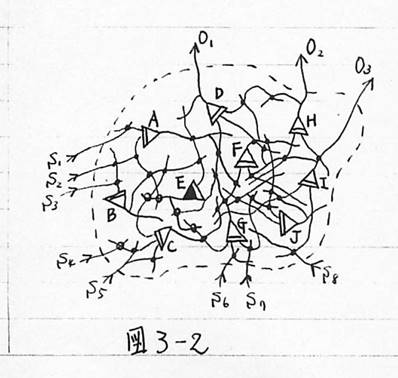
図3-2 神経回路網
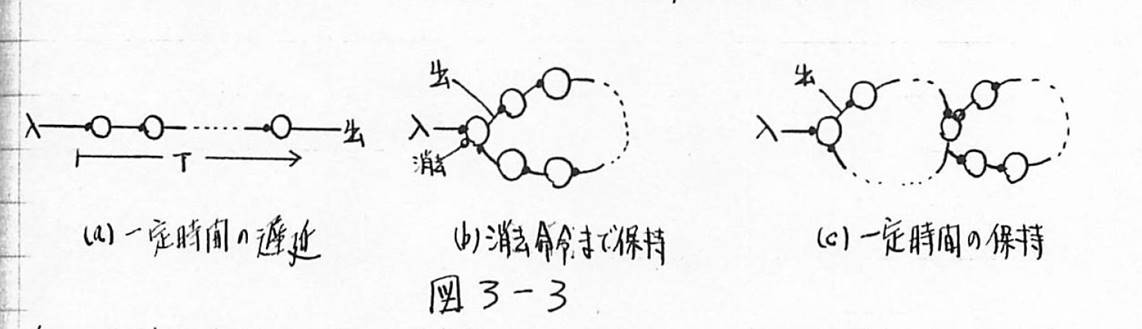
図3-3 神経細胞の直列接続による時間の表現
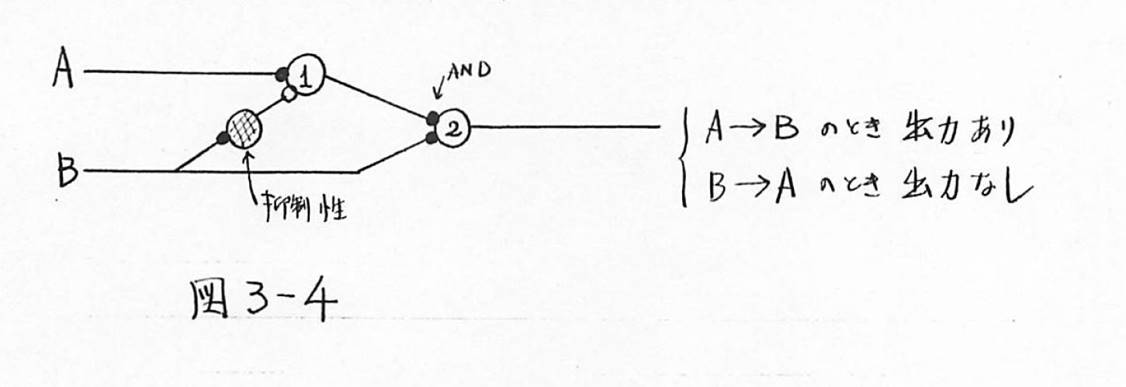
図3-4 神経細胞の直列接続による順序の表現
3.2 組み合わせモデル
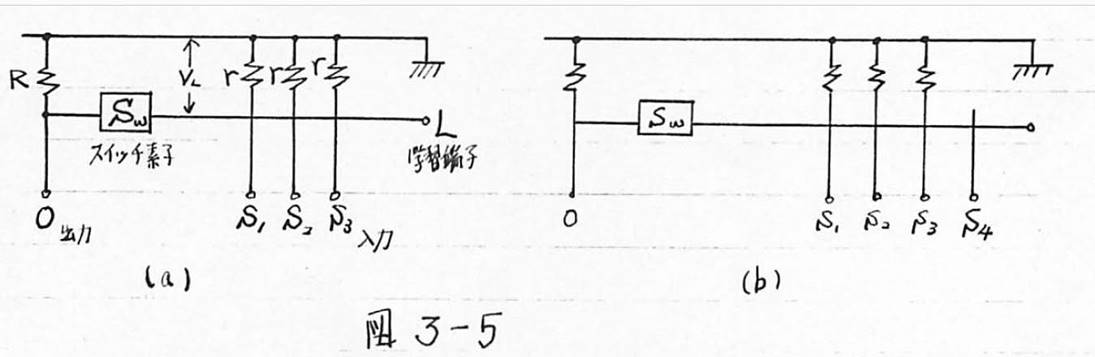
図3-5 (a)興奮性シナプスのAND回路表現 (b)抑制性シナプスの回路表現
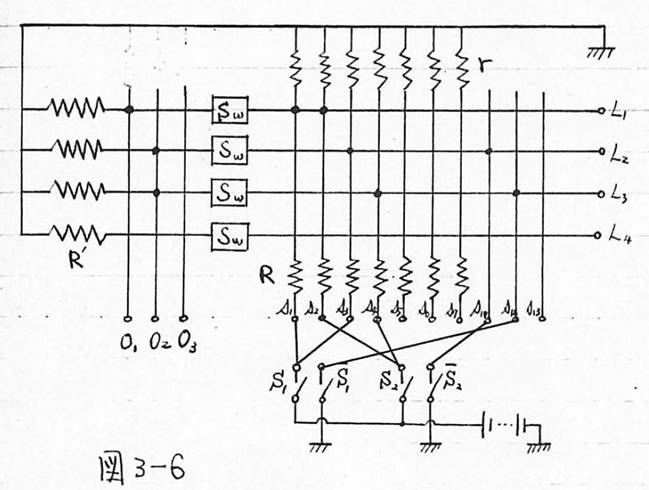
図3-6 脳のモデルの回路表現
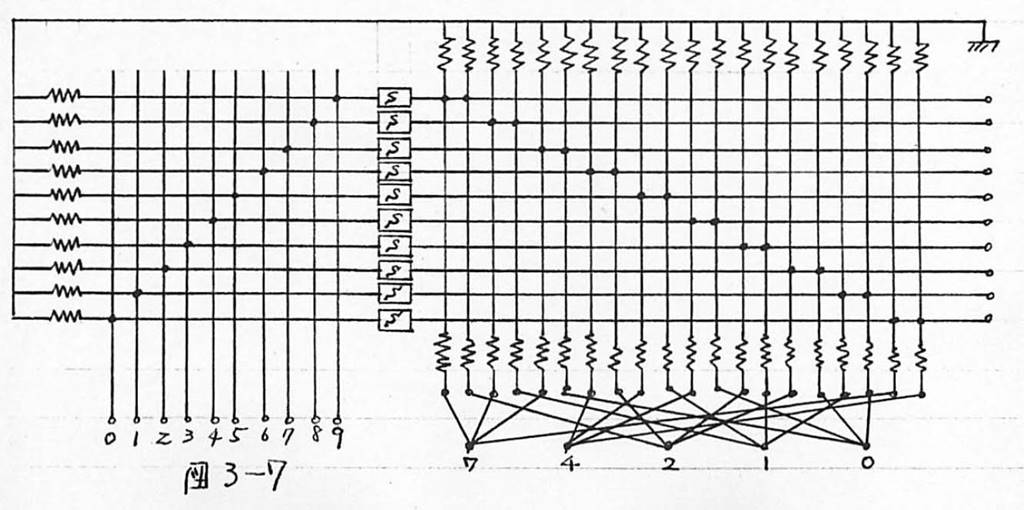
図3-7 例1:符号変換回路
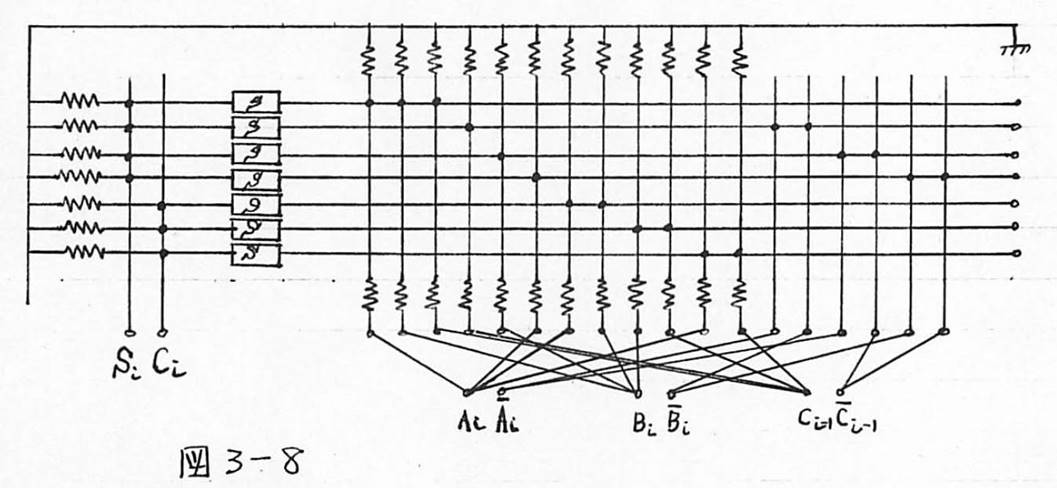
図3-8 例2:加算器
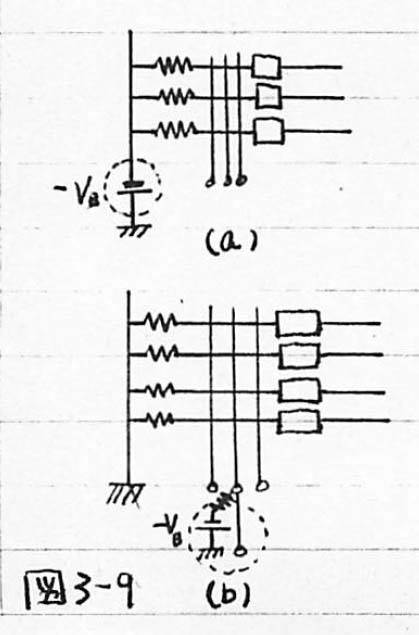
図3-9 応用1:入力端子数と入力数の比による動作
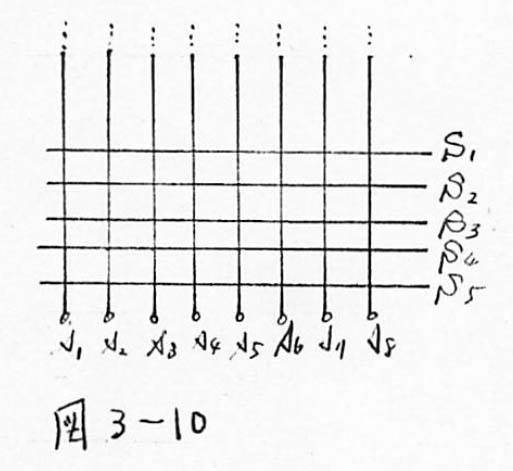
図3-10 応用2:格子状の配線の短絡を利用した配線の単純化
3.3 集積回路化について
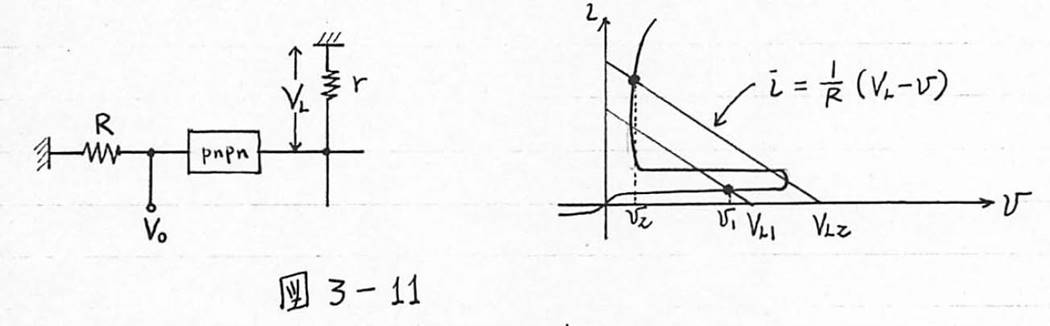
図3-11 スイッチ素子にpnpnダイオード利用の例
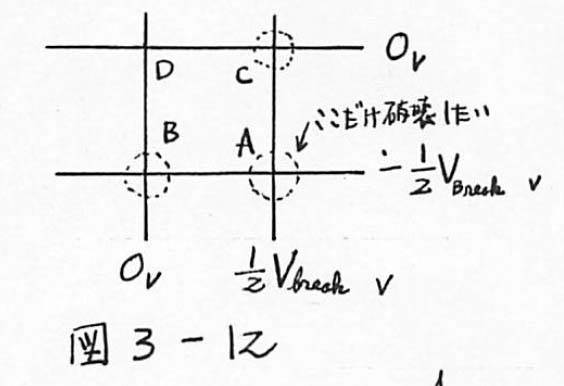
図3-12 SiO2 の絶縁破壊を利用の場合
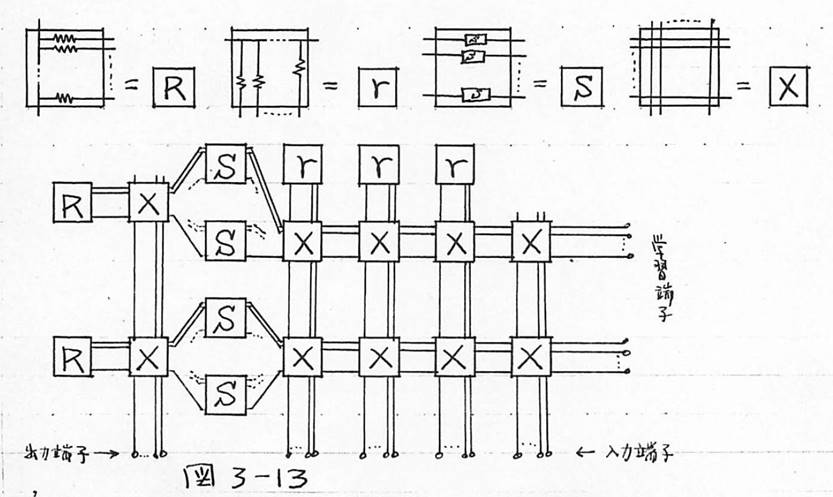
図3-13 基本素子のIC化のよる大容量化
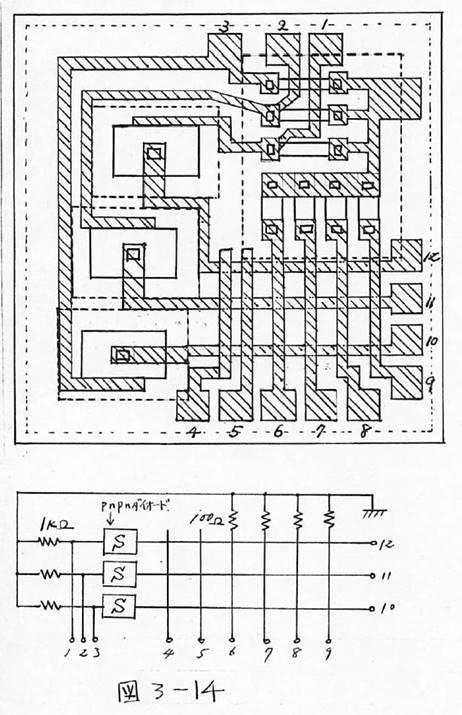
図3-14 半導体集積回路の設計
3.4 学習機械としての評価
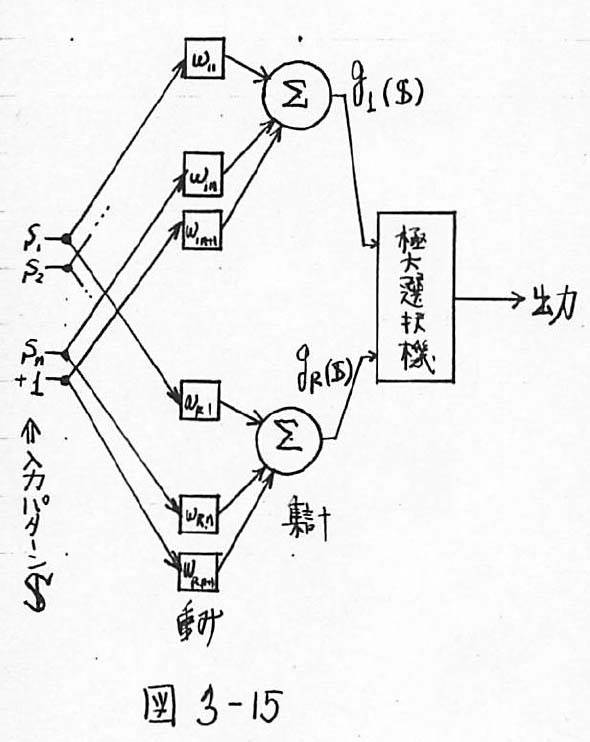
図3-15 学習機械の例:線形機械の重み調整による極大選択機
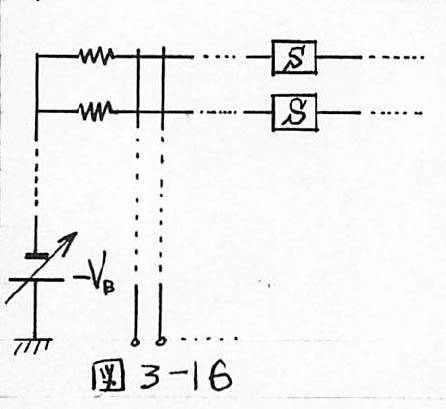
図3-16 提案システムによる極大選択機の実現
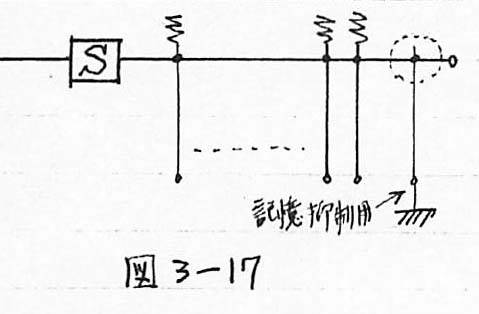
図3-17 提案システムにおける記憶の消去の実現
第4章 結言
この論文では、第1章では、条件反射の解析ということを念頭に置きながら、それに役立つと思われる生理学的な資料を再編成してみた。その過程で、巨視的な事実としての条件反射活動と微視的な事実としての神経細胞の働きとを、如何にして一直線上に捕らえることができるかということ、即ち、1.2で考えた反射活動における脳のシステム(図1-3)を神経細胞がどのようにして構成していくかということが、非常に大きな問題として残った。特に、条件反射の形成過程で、神経細胞が如何なる機能を果たしているかという問題は、そこに学習機能の本質が横たわっていると思われるので、いっそう重大である。
そこで、第2章では、生理学的な事実の助けを借りながら、且つ、それに制約されながら、その問題の解決を試みた。この分野では、Harth, Caianiello等の試みがあり、まず彼等のモデルの検討から始めた。その結果、学習効果として、興奮した神経細胞の間の結合係数の増加を考える方法は、結合係数に初期値を与える必要のあることから、出発点の影響が大きく、丁度、皮質を除去した動物のような学習性を持ってしまうことがわかった。そこで、私は、「自由な神経細胞」を提起し、これを新皮質における条件反射形成の主役と考え、条件反射モデルには、この自由な神経細胞の機能を持たせる必要があるという結論に達した。
次に第3章では、その結論に従って、条件反射をする脳のモデルを考えた。ここで最も大きな問題は、この自由な神経細胞にどのような学習効果を与えるかということである。これについては、3.1で述べたように、神経細胞の機能を従来のように閾値素子と考えず、AND回路のOR回路として考えたことに関連して、ひとつの試みとして、2.4の終わりに示したように、自由な神経細胞は同時に興奮したすべての神経細胞と結合してAND回路を形成すると仮定した。その結果、図3-6に示すような条件反射を行う脳のモデルが出来上がった。
このモデルを学習機械として考えたとき、記憶に絶縁破壊を用いたため、応用面での柔軟性に欠けるような印象を受けやすいが、3.4でも示したように、例えば、出力側の接地面に負のバイアスをかけるというような工夫をすれば、いろいろな機能を備えうるので、入力パターンが2値論理パターンである限り、幅広い応用の可能性を秘めている。ただ、時間的制約があって、ほとんどその方面の研究をやれなかったので、工学的応用分野の研究や、他の学習機械との比較検討は今後の課題として残されている。
謝辞
最後に、末筆ながら、親切にご指導くださった斎藤正雄先生をはじめ、有意義な討論をされた斎藤研究室の方々、そして、二度にわたり、快くご意見をくださった医学部の矢島助手に感謝します。
また、同じ分野を志し、常に良き相談相手であった友人伊藤憲治君に感謝する。
文献
[生理学、生体工学関係]
1.時実ほか:脳と神経系、岩波講座、現代の生物学、1966.
2.島崎ほか:脳の働き、共立出版、1966.
3.A.B.コーガン:脳生理学の基礎、川村訳、岩波書店、1963(原 1959).
4.エヌ・タラーウゴット、エリ・バローノフ&ア・リーチコ、人間の大脳活動、
樺島ほか訳、世界書院、1962(原 1957).
5.イェ・エヌ・ソコーロフ:知覚と条件反射、金子ほか訳、世界書院、1965(原 1958).
6.ア・オ・ドーリン:ノイローゼの生理学、岡田訳、世界書院、1966(原 1962).
7.梅岡ほか、学習心理学、誠信書房、1966.
8.E.M.Harth:"Brain Models and Thought Process" Automata Theory, Academic Press, 1966.
9.E.M.Harth and S.L.Edger: Association by Synaptic Facilitation in Highly Damped
Neural Nets, Biophisical Journal, 7, pp.687-717, 1967.
10.E. R. Caianiello : Outline of a theory of thought-process and thinking machine,
Journal of theoretical biology, 2, 204-235, 1961.
11.S.Deutsch : Models of the nervous system, John wieley & sons, Inc., 1967.
12.K.S.Abuladze : Central inhibition of reflexes and the problem of the coupled
activity of cerebral hemispheres, Brain reflexes, Progress in brain research,
volume 22, Elsevier publishing company, 1967.
13.E.A.Asratyan : Some peculiarities of formation, functioning and inhibition of
conditioned reflexes with two-way connections, Brain reflexes,
Progress in brain research, volume 22, Elsevier publishing company, 1967.
14.S.A.Sarkisov : The structure and functions of the brain, Indiana university press,
1966.
15.D. O. Hebb : A textbook of psychology, W.B.Saunders company, 1958.
(白井ほか訳、行動学入門、紀伊国屋)
16.山口登:思考過程のモデル、東大大学院電子論文輪講資料、1968.
17.浜ほか:神経の生物物理、生物物理学講座、吉岡書店、1966.
18.L.D.Harmon & E.R.Lewis : Neural modeling, 1966, Physical rev. 46:513.
19.H. W. Magoun : The waking brain, 2nd ed., C.Thomas Illinois, 1962
(時実訳、脳の働き、改訂新版、朝倉書店、1967)
20.N. Wiener : Cybernetics, 2nd ed., The MIT press, Mass., 1961.
(池原ほか訳、サイバネティックス、第二版、岩波書店、1962)
21.デカルト:方法序説、小場瀬訳、角川文庫、1963(原作1637)
22.ド・ラ・メトリ:人間機械論、杉訳、岩波文庫、1932(原作1747)
23.エンゲルス:自然の弁証法、田辺訳、岩波文庫、1956(原作1876)
[オートマトン、学習機械関係]
24.オートマトン特集号、電子通信学会誌、1963.11、46巻、11号
25.N. J. Nilsson : Learning machine, Mc-Graw Hill, 1965.
26.A.M.Uttley, Conditional probability machines and conditioned reflexes,
Automata studies, Princeton Univ. press, 1956.
27.W.A.Ainsworth, On the efficiency of learning machines, IEEE trans. SSC-3, 2,
pp.111-116, Nov. 1967.
28.坂井利之:パターン認識の理論、共立出版、1967.
29.室賀三郎:オートマトン入門、共立出版、1960.
30.喜安:デジタル回路の数学、共立出版、1960.
31.北川敏男:学習実験、共立出版、1966.
32.北川敏男:学習制御および学習制御機械、共立出版、1966.
33.大照、加藤、竹谷:Stochastic learning machine, 信学会 オートマトン研究会資料、
A68-4,1968-04.
34.木村、石井、本田:閾素子によるパターン認識、信学会、オートマトン研究会資料、1964-6.
35.磯道、猪俣、飯島:パターンの識別理論、オートマトン研究会資料、1966-1.
36.北橋、手塚、笠原:多数決論理回路における入力集合のSeparabilityについて、信学会、
オートマトン研究会資料、1966-1.
37.藤崎、倉田:パターン認識における評価およびパラメーター選択に関する一考察、信学会、
オートマトン研究会資料、1964-5.
38.戸田、伊達:多数決素子の適応的構成、信学会、オートマトン研究会資料、1964-5.
39.鈴木、牟田:パターン認識におけるパラメーター評価、信学会、オートマトン研究会資料、
1964-5.
40.玄地、森:従属性を考慮した特徴評価法、信学会、オートマトン研究会資料、1964-5.
41.宮本友義:記憶、信学会、オートマトン研究会資料、1968-09.
[集積回路関係]
42.小田川、中川,垂井:集積回路とその応用、日刊工業新聞社、1968.
43.集積回路ハンドブック、集積回路ハンドブック編集委員会、丸善、1968.
44.R.M.Warner, Integrated circuits, Motorola Inc., 1965.
(小田川ほか訳、集積回路、近代科学社)
45.P.Richman, Characteristics and operation of Mos field-effect devoce,
Mc-Graw Hill Inc.,1967.
(斎藤ほか訳、MOS電界効果素子、近代科学社、1968)